|
|
|
Inside
Series
Site Search
New book:
Starting Science from God.
|
Chance, Evolution and the New Wordby Charlotte Gyllenhaal-Davis Although evolutionary theory is often used in New Church literature as an example of the grossest of materialistic thought, it is still an intriguing concept to many in the Church. Since we are acquainted with the spiritual sense of the Word, we are not bound to the Creation Story in Genesis for the history of the natural world. We are all aware of the diversity of extinct plants and animals depicted in the fossil record, and of the fact that humans have been on this earth for a relatively short time. The possibility of some sort of ascent to man is often acknowledged as valid. Johannes Klotter [2] creatively attempted to build a New Church evolutionary theory using ideas from Swedenborg, but postulated entities and processes beyond what seems necessary to a biologist. With a proper understanding of both New Church doctrine and evolutionary theory, however, we need not search for a set of observations, entities, or physical theories different from those used by contemporary science. The more recent developments of evolutionary and genetic theory give a coherent explanation of the process of change in the fossil record, including its apparent gaps and jumps. If we keep in mind the teachings of the Writings concerning chance and influx, modern evolutionary theory becomes eminently compatible with New Church philosophy, and thus takes its appropriate position in our minds as a plausible model of the workings of Divine Providence in the ultimate natural plane. What is Evolution? Why have biologists developed evolutionary theory? It springs from three basic observations: the presence in the fossil record of life forms that are extinct today, the ability of breeders to alter the behavior and appearance of domestic plants and animals through artificial selection, and the obvious physical similarities or affinities among certain organisms. The fossil record is both incomplete and amenable to various human interpretations. Sometimes it shows a succession of gradually changing life forms, and sometimes new forms arrive on the scene relatively suddenly. Sudden arrivals have been used by those advocating fiat creation as evidence of instantaneous Divine formation of new species. Early Darwinians concentrated on the instances of gradual change. Current evolutionary theory can explain both types of change in an orderly way. The fiat creationists and Darwinians usually agree about the implications of plant and animal breeding. Natural selection, which Darwin discovered partly through studying the variation in domestic plants and animals, can be observed today. It is a widely misunderstood process; its capsule form, "the survival of the fittest," is often wrongly associated with "nature red in tooth and claw." The combatative model of natural selection, in which the strongest of a species kill the weakest and thus survive, is no longer considered valid. What actually survive in natural selection are the fittest genes, or those which confer upon an organism bearing them a reproductive advantage relative to other members of its species. Those organisms that bear more and better adapted offspring are more likely to have their genes represented in the next generation than those organisms that bear poorly adapted offspring which perhaps die before they can reproduce. Through natural selection, the genetic background of a species responds to the pressures of its environment, and deleterious genes gradually fade away. In the Darwinian synthesis, natural selection leads to speciation (the formation of new species) when it acts upon populations that move into new niches and habitats. The resulting species bear some resemblance to their parent species, which allows taxonomists to construct "evolutionary trees," indicating ancestral and descendant species. Modern Neo-Darwinian evolutionary theory, however, recognizes several mechanisms of evolution besides natural selection; I shall explain some after examining the concept of chance and its role in evolution. Chance What is the biological concept of chance, or randomness, as it is usually referred to in science? A random process is characterized by the fact that we cannot predict in advance what the result of the process will be for an individual case, but only say that each particular result has a certain probability. We can predict that for a group of cases, so many will have one result, so many a second result, so many a third result. Our predictions are based on mathematical descriptions of the logical or previously observed outcomes of the process. One familiar example of a random process is coin tossing. In coin tossing, there are two possible results: heads and tails. In a natural or random situation, we know that the two results are equally probable: each has a probability of 50% or .50. By a mathematical expression, the binomial distribution function, we find that if we toss a coin twice, the probability of getting two heads is 2570 or .25, the probability of two tails is .25, and the probability of getting one heads and one tails is .50. We can calculate the probabilities of getting a certain proportion of heads and tails for any number of coin tosses we choose. As we toss the coin more times, we find it becoming progressively more likely that we will get equal numbers of heads and tails, and less likely that we will get very different numbers. If in a series of 100 coin tosses, 80 result in heads and only 20 in tails - a low-probability occurrence - we suspect that normal probability is being disrupted, or that our coin is weighted.
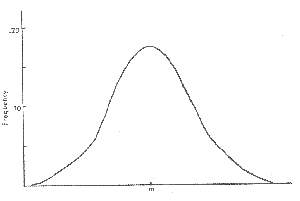
Figure 1. The normal curve A familiar example of a random process is pictured by the bell-shaped, or "normal" curve seen in Figure 1. The normal curve describes characteristics of populations which vary continuously, such as height or weight, rather than having two or more discrete states as in the previous example. It is deduced mathematically by assuming that there are many factors influencing the quantity we are studying (e.g., several genes, supplies of various vitamins, exposure to several environmental factors), each of which acts as if it were a character with two or more equally probable states, and could thus be modeled by the binomial distribution. The mathematical derivation for the probability of occurrence of all the possible values of the characteristic of interest yields the normal curve, in which those values in the middle of the range occur much more frequently than those at either end.[3] The value exactly in the middle ("m" in Figure 1) is equal, to the average, or arithmetic mean of all possible values.
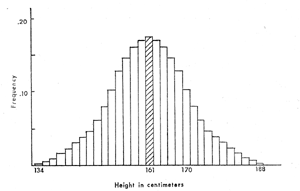
Figure 2. Idealized bar graph or histogram of the heights of women aged 35 to 44 The normal curve describes accurately the variation of many natural phenomena. If we measure the height in centimeters (cm) of a large number of women and count how many are 160 cm tall, how many 161 cm, and so on for the entire range of heights, we can make a bar graph, illustrated in Figure 2.[4] The most frequently occurring height, 161 cm, or the mean is shaded. The least frequently occurring heights are 134 cm and 188 cm, in this graph, and are at either end of the bar graph. Heights become less and less frequent, or less probable as one moves out to either end of the bar graph. The rate at which the frequencies drop off is specified in a normal curve, and we are able to say that 33% of the population falls between the mean height and a particular taller height (170 cm for this graph). We can also say that if we take a new individual, there is a 33% chance or probability that her height falls between 161 cm and 170 cm. In fact, we can calculate the probability that her height will fall between any two heights. Height is not the only characteristic of a population that can be described in this fashion; and the normal curve is not the only one that describes random variation of characteristics in populations. Another important feature of random processes is that they repeat themselves. If we do an experiment consisting of tossing a coin 100 times, we find the cumulative observed frequency of heads approaches .50 more closely as we reach the hundredth toss. Consider an imaginary world, however, where a series of coin-tossing experiments do not result in probabilities approaching .50 each time. One time the probability of heads might approach .67, the next time, .81, the next time .25. In this world, we could not predict probabilities. If all events in this world resulted in unpredictable and unstable probabilities, we could call it chaotic. Contrast the orderliness of our actual universe with the jumble of the alternate chaotic world: the utility of our statistical theory depends on the stability of our probabilistic world. To summarize, science sees chance as an orderly, workable phenomenon. Mathematical descriptions of the outcomes of random processes also describe characteristics of the natural world. These descriptions can be used to make probabilistic predictions of the results of natural processes which are regularly borne out when the mathematics is correctly applied. What do the Writings teach about chance? They make it quite clear that chance (or fortune) is "Divine Providence in ultimates."[5] Divine Providence in ultimates, in the "least things" that operate in the formation of observable natural quantities, thus would seem to act in ways that are amenable to orderly probabilistic description. We can see neither the means nor the ends of these least events, and they are quite beyond our influence, unless we attempt to disrupt natural processes and inhibit the free operation of chance. Providence in ultimates, to our eyes, is probability. Providence in the least things of nature acts as it does in the greater. It leaves people in freedom to accept or reject the Divine invitation as they choose; many have used chance to justify their rejection. Random processes cannot be predicted in advance for individual cases, but hindsight - the binomial distribution function and the normal curve - can perceive the results after the process is over. And the orderliness, the repeatability of our observations on the trends of random processes, means that we live in a universe in which we can use our rationality. The application of human intelligence to random processes and the prediction of probabilities is at the heart of modern scientific and technological progress. Chance and Reproduction What has chance, or Providence in ultimates, to do with natural selection and other modes of evolution? Natural selection results from the differing capacities of individuals of the same species to reproduce. The initial processes in sexual reproduction are chance processes. In sexual reproduction the union of male and female gametes, sperm and egg, produces a zygote, which grows into a fetus and emerges as a new individual at birth. The two gametes carry all the genes that will control the development and appearance of that individual. How are gametes formed? Each cell of our body has 46 chromosomes - 23 pairs of chromosomes. Other species have more or fewer chromosomes. These chromosomes contain deoxyribonucleic acid (DNA), which makes up genes; we can have two alleles, or alternate forms of a gene, one on each member of a pair of chromosomes. In gamete formation, the cell that will become sperm or egg cells undergoes meiosis, or reduction division. The chromosomes line up in the middle of the cell by pairs, and the pairs split up, each member migrating to one end of the cell. The cell then has two groups of 23 chromosomes. A cell membrane forms between the two groups, and the cell splits into new cells which become gametes. Each gamete then has only 23 chromosomes, one of each pair. It also has only one allele of each gene. Reduction division entails chance when the chromosome pairs split up and migrate to opposite ends of the cell. Exactly what causes one member of a pair to migrate to one pole of the cell, the other to the opposite pole, we do not know. We do know that chromosome migration is a random process: a chromosome usually has an equal probability of moving to either end pole. Occasionally, a gene will mutate in a way that can distort this process, but there is rapid natural selection for the rest of the genes to suppress these "segregation distorters" (such suppressive interactions among genes at the cellular level are quite common). The "fairness" of meiosis is of great importance to every species, since without random separation of alleles, the genetic variability which allows a species to adapt to changing environments would be restricted.[6] At conception, as well, the choice of exactly which sperm and which egg unite is random one, that is, one controlled by Divine Providence in ultimates in a probabilistic way. This is confirmed by the fact that, if we know which alleles for a particular genetic trait each parent has, we can predict the probabilities that their children will have each of the possible combinations of alleles using probability theory - though we cannot predict whether an individual child will or will not have a particular genetic combination. Random combinations of sperm and eggs also help ensure that the variation mentioned above will be maximal. The importance of random separation of chromosomes at meiosis, and the random nature of gamete conjunction, indicate that these basic reproductive processes are controlled by Divine Providence working in ultimates. The results of these chance processes, and of environmental influences that later affect the individual's development, are manifested in populations whose physical or behavioral variation is described by the binomial distribution function, the normal curve, or other probability distributions. Why is it providentially important that such variation exist? On a spiritual level, we might speculate that the variation promotes "the image of the Infinite and the Eternal in the variety o£ all things,"[7] or fulfills the requirement that "there must be variation in everything from its greatest to its least," [8] since good, truth, or beauty can be perceived only in comparison to what is less good, less true, less beautiful, even down to what is evil, false, or ugly. More practically, variation such as that pictured in the normal curve provides genetic flexibility for species that confront changing environments. In the normal environment, those individuals whose characteristics fall at or near the mean value for the species usually reproduce most efficiently. In a different environment (hotter, colder, or wetter, for instance), individuals near the mean might reproduce poorly, whereas those at one end of the normal curve might reproduce quite well. Genetic variability thus enables a species to survive in an expanded range of environments, or to persist in its range during times of climatic change. Chance and Evolution Divine Providence in ultimates works at many levels in current evolutionary theory. Its effects begin at the molecular level, with the random aspects of genetic mutation. In mutation, one allele turns into a second allele, due to factors of which we are not completely sure, but which include x-rays, cosmic rays, and various chemicals. The process involves the substitution of as little as one deoxyribonucleotide (component of a DNA molecule) for another. The allele may turn from a dominant form to recessive form, or a recessive to a dominant form, or from one co-dominant form to another. Each gene has a certain probability of mutation; a transition in one direction may be more probable than the reverse mutation. We can predict the probabilities of mutation from observation - our knowledge of molecular genetics is insufficient to allow us to predict mutation rates based on the configurational stability of various forms of DNA. But, as always when working with Providence in ultimates, although we can predict the probability of mutation, or even raise it by exposing organisms to x-rays or other mutagens, we cannot absolutely predict whether a particular individual's gene will mutate. Given both the variation induced by meiosis and the presence of mutations that can alter an individual's fitness, natural selection becomes inevitable. If all the possibilities on the normal curve are to be filled, some individuals will naturally be healthier, more sociable, or otherwise better reproducers than others. Because deleterious mutations that inhibit reproduction - and most mutations are deleterious - are weeded out during natural selection, the adaptedness of each species gradually improves. Could this be an image in nature of the Lord's gentle, upward leading of man's spiritual life? If natural selection results in gradual change, how do we explain events in the fossil record that indicate rapid change? [9] Current evolutionary theory has several descriptions, including processes that involve large populations and cause great morphological changes in response to slight environmental changes, as well as processes that occur in small populations evolving at the edge of a species' range in unusual environments. The former processes depend on a property of genetic systems known as "canalization," or genetic homeostasis. Canalized traits arise from groups of genes that interact to modify the effects of other genes, some of whose alleles may be incompatible with the proper development of the trait. In addition, canalization gives a species greater environmental flexibility, as canalized traits will develop properly in a variety of different environments.[10] We do not understand the mechanisms of canalization; the diversity of genetic functioning has but lately been realized. In addition to interacting groups of genes, a number of single genes have been discovered which regulate the activity of other genes. Scientists have hypothesized that these regulatory genes are extremely important in the individual's development, since they act as "timers" that "turn on" and "turn off" genes at critical developmental stages. Regulatory genes could logically be involved in canalization as well. The genetic system is homeostatic only within certain limits. Dodson and Hallam have recently theorized that when a species expands its environmental range until it reaches the threshold at which its original canalization can no longer function, it undergoes unusually intense natural selection until a new homeostasis is reached.[11] This hypothesis is based on the mathematics of catastrophe theory. If the proper genes are affected, a very different-looking organism can appear quickly and with relatively little genetic change. It is interesting to note here that the amino acid chains (protein components whose synthesis is directed by genes) of humans and chimpanzees are more than 99% similar, implying that most of their genetic endowment is very similar as well.[12] The vast differences in appearance and behavior of the two species suggest that the crucial regulatory genes, which probably control development, alter the mode of expression of other genes, and may be involved in canalization, might be the ones that actually differ in humans and chimpanzees. Regulatory genes mutate randomly, as do other mutations in the population, and could be viewed as a manifestation of Providence working in ultimates. Natural selection acting on regulatory genes at the environmental limits of genetic homeostasis may work best on large populations. But founder effect and random genetic drift promote speciation only in small, isolated populations in habitats that are marginal or newly available to a species. Founder effect describes the observation that when a small group is isolated from the rest of its species, the genetic composition of its founders will largely determine that of its descendants. If the founders possess some rare gene, deleterious in the usual environment of the species, the descendants probably will also possess that gene. It will be more common in the descendant than in the original population, and since there will be fewer reproductive competitors, natural selection against the deleterious gene will be less severe. It seems obvious that the Providential hand might select the founders of a new population. An example of founder effect is the Ellis-van Creveld syndrome, a type of genetic dwarfism known only in a group of Old Order Amish living in eastern Pennsylvania. In 1939, this form of dwarfism was distinguished from dwarfism found among Amish living in other areas of the United States. All the individuals affected by it were found to be descendants of one couple who settled in eastern Pennsylvania in 1744, suggesting that couple was the source of the gene concerned. Because the Old Order Amish do not marry outside their faith, and have apparently migrated only slowly from settlement to settlement, the descendants of the founding couple and their neighbors formed a small, isolated population. The presence of the gene causing Ellis-van Creveld syndrome in the founders of the population and its persistence through several generations of isolated inbreeding are common features of situations involving founder effect. Random genetic drift is another consequence of the random assortment of chromosomes in meiosis. Each gamete carries only half the chromosomes, thus only half the allelic forms that its parent possesses. If a couple has only one child, only half of the alleles of each parent will be represented in the next generation. If they have more children, more of their alleles will be represented, but they would need to have a great many children indeed to make sure every allele that both parents carried would be present in the next generation. In a small population in which each pair raises only a few offspring, it is likely that several chromosomes in each generation will not be included in the gametes that succeed in forming zygotes and becoming new individuals. Some allelic forms will probably be lost in every generation. Which alleles are lost is determined by chance, that is, by Divine Providence in ultimates. Other alleles will increase in frequency due to chance. Genetic "drift" refers to the resulting changes in proportions of the different allelic forms in each generation. The randomness of genetic drift originates in the probabilistic nature of meiois. Founder effect and random genetic drift can both lead rather rapidly to genetic and morphological change in a species. They could work even more rapidly if the alleles that increased or decreased in the population were forms of regulatory genes. If a new species developed in one of these ways - or, more likely, all of them acting together - it would develop rapidly. If the new species developed in a small, isolated group, it seems unlikely that we would happen to find intermediate forms in the fossil record. But, if it developed into a better competitor than its parent species, it might reinvade the parental habitat, rapidly increase in numbers, and out-compete the original species.[13] Thus, sudden new arrivals in the fossil record might represent migration and competition rather than untoward gaps in the fossil record, or the sort of Divine creation from nothing that fundamentalists' interpretation of the Word seem to demand. That the new species was a Divine creation is undoubtable ; the possibilities for Providence working in the ultimates of nature are numerous, as attested by the several random processes we have examined. Could these processes work in human evolution? I see no reason why they should not. Human evolution is a complex subject, and we do not know at what point our primate line became receptive to the human soul, rather than a generalized "primate soul." Nor do we know what primate was the direct ancestor of humans. The genetic similarities of humans and chimpanzees lead us to suspect that both of these species evolved from a common animal ancestor which was fairly close to both in genetic make-up. We might hypothesize that relatively few mutations in regulatory genes, particularly those controlling development of the nervous system, could effect the change from a form receptive of an animal soul to one receptive of a human soul. If carriers of the crucial regulatory genes were Providentially isolated and given a chance to reproduce, the change in the homeostatic genetic system could happen quickly and decisively, through any or all other genetic mechanisms discussed above. Dr. G. R. Doering describes some of the spiritual and natural details of the way in which the first human souls might be implanted in receptive primates.[14] Humans have probably been modified as to details of physical appearance and stature, and possibly even modified to become more fully receptive of human souls since the first humans lived on earth. I feel, though, that there is no reason to suppose that the natural human body could not have been created by the mechanisms described in modern evolutionary theory. The concept of chance as the operation of Divine Providence in least things allows the New Church person the ability to live with equanimity in the probabilistic universe. In practice, one's dealings with chance differ little from those of nonbelievers; it is simply that we acknowledge a Source of natural variability, and the forms that it takes, rather than taking them as given. The results of chance events and the ability to predict where chains of such events will lead are as far beyond our knowledge as anyone else's. Had the first humans had our statistical knowledge, they would have had as little idea of the ways in which chance and natural selection would alter their bodies and brains as they had without that knowledge. They might have felt, however, that wherever the operations of chance might lead, the consequences would be Divinely presented for them to respond to freely and in a way that would help them - already images of God - to become true likenesses of God. Footnotes 1. I would like to acknowledge E. G. Leigh, Jr., for his stimulating thinking on evolution; Todd McLin Davis and Grant R. Doering for their most helpful comments; and Pat Probst for her typing assistance and comments; any questionable interpretations, however, are my responsibility and not theirs. Fellow scientists should note that the biological and statistical explanations are aimed at nonscientists, and not intended to be an exhaustive treatment. 2. Klotter, J., 1975. "Evolution and the Origin of Man." THE NEW PHILOSOPHY, 78: 171 3. Sokal, R. R., and F. J. Rohlf. 1969. Biometry. (Freeman, San Francisco), p. 102. 4. The bar graph is made by counting heights in groups with a range of three centimeters (e.g. 160, 161, and 162 centimeter heights are grouped), which is a little over an inch. The mean height, 161 centimeters, is about 64 inches or 5 feet, 4 inches. Data for the graph are from: Altman, P. L., D. S. Dittmer, 1972. Biology Data Book, 2nd edition. Vol. 1, p. 201 5. DP 212, AC 6493, 6494. 6. Leigh, E. G., Jr., 1977. "How does selection reconcile individual advantage with the good of the group?" Proc. Nat. Acad. Sci. U.S.A. 74: 4542-4546. 7. DP 56. 8. DP 24. 9. Eldredge, N., and Gould, S. J., 1972. "Punctuated equilibria: an alternative to phyletic gradualism." In Models in Paleobiology, ed. Schopf, T. J. M. (Freeman, Cooper; San Francisco) pp. 83-115. 10. Waddington, C. H., 1942. "Canalization of development and the inheritance of acquired characters." Nature 150: 563. 11. Dodson, M. M., and A. Hallam, 1977. "Allopatric speciation and the fold catastrophe." American Naturalist 111, pp. 415-433. 12. King, M. C., and A. C. Wilson, 1975. Science 188: 107. l3. Gould, S. J., 1977. "Evolution's erratic pace." Natural History 86 (5) : 12-16. 14. Doering, G. R., 1968. "Evolution." THE NEW PHILOSOPHY 71: 68-77. -The New Philosophy 1978;271-283 |
Up
|
|
|